


La maladie corticale du hêtre
Dans cette page :
- Historique
- Mécanismes d’infection
- Cycle biologique de l’insecte
- Cycle biologique de la maladie et symptômes
- Évolution de la maladie et distribution
- Dégradation des bois et impacts
- Impact sur la dynamique forestière
- Recommandations d’aménagement
Historique
La maladie corticale du hêtre (MCH) résulte d’une interaction entre un insecte, la cochenille du hêtre (Cryptococcus fagisuga Lindinger) et les deux champignons pathogènes suivants : 1) Neonectria faginata (Lohman et al.) Castl. et Rossman (espèce exotique) et 2) Neonectria ditissima (Tul. et C. Tul.) Samuels et Rossman (espèce indigène). N. faginata n’existe que sur le hêtre à grandes feuilles, Fagus grandifolia Ehrh, tandis que N. ditissima, appelé aussi le chancre nectrien, infecte plusieurs espèces de feuillus. La cochenille du hêtre et N. faginata ont été introduits accidentellement près du port de Halifax en Nouvelle-Écosse en 1890 par l’importation de plants de hêtre pourpre, Fagus sylvatica f. purpurea (Aiton) C.K. Schneid, provenant de l’Europe. Ce n’est qu’en 1911 que les premiers symptômes de la maladie ont été détectés dans les forêts naturelles. Par la suite, l’insecte et la maladie se sont propagés vers le sud-ouest et le nord-ouest du continent pour s’étendre dans les provinces canadiennes des maritimes et certains États américains. La MCH a été découverte pour la première fois au Québec près du lac Témiscouata, en 1965, et en Ontario en 1999. La maladie s’est propagée à un rythme moyen estimé de 16 km par année pour couvrir de nos jours quatorze États, comprenant la Nouvelle-Angleterre, puis s’est étendue de la Caroline du Nord jusqu’au Michigan, et dans cinq provinces canadiennes comprenant le Québec, l’Ontario, le Nouveau-Brunswick, la Nouvelle-Écosse et l’Île-du-Prince-Édouard.
Cette maladie a un impact considérable sur son hôte, le hêtre à grandes feuilles, et par conséquent sur la dynamique des peuplements forestiers.
Mécanismes d’infection

Photo 1 – Cire blanche à l’aspect laineux secrétée par la cochenille du hêtre (Photo : N. Nadeau-Thibodeau)
La maladie survient lorsque les spores des champignons s’introduisent par des blessures faites à l’écorce, entre autres celles causées par la cochenille du hêtre. Ces insectes suceurs créent des milliers de microblessures dans l’écorce afin de se nourrir. Une substance produite par la cochenille empêcherait la cicatrisation des blessures. Les spores des champignons pathogènes peuvent y germer, et ce, sans réaction immédiate de l’arbre.
La cochenille filamenteuse, Xylococculus betulae (Pergande), peut aussi s’attaquer au hêtre et ses dégâts peuvent précéder ceux de la cochenille du hêtre et ainsi contribuer à l’introduction des champignons. Bien que la maladie soit le plus souvent associée à la présence de ces insectes, les stress climatiques, y compris les épisodes de sécheresse estivale, les températures élevées et le froid extrême, peuvent rendre l’arbre sensible aux infections de Neonectria. Les cochenilles ne sont donc pas une condition préalable à l’infection par les espèces de Neonectria.
Cycle biologique de l’insecte
La cochenille du hêtre mesure environ 1 mm de longueur à maturité. Elle est de forme elliptique de couleur jaune avec de minuscules yeux brun rougeâtre. Elle sécrète une cire blanche, et s’en couvre progressivement, laquelle cire a un aspect laineux, signe caractéristique de la présence de l’insecte à la surface du tronc des hêtres (photo 1). La reproduction de la cochenille est parthénogénétique, ce qui signifie que la population n’est constituée que d’individus femelles. Les femelles apodes (immobiles) déposent leurs œufs dans les stries de l’écorce de la fin de juin au début d’août. Une fois écloses, les larves munies de six pattes peuvent rester sur place près de la femelle, se déplacer sur le tronc, voire migrer sur un arbre voisin à l’automne pour former d’autres colonies. Seul l’insecte immature peut se mouvoir. Plus tard, les jeunes larves vont insérer leur stylet dans l’écorce. Elles perdront alors leurs pattes pour devenir des femelles immatures et immobiles, puis elles hiverneront pour devenir matures. C’est au printemps suivant qu’à leur tour elles pondront leurs œufs.
La cochenille filamenteuse s’attaque non seulement au hêtre, mais aussi à d’autres essences telles que le bouleau à papier et le bouleau jaune. Elle s’enfonce dans l’écorce produisant ainsi une blessure rappelant un petit chancre causé par la MCH. Avec le temps, l’écorce fendille perpendiculairement pour former une petite croix (photo 2). Il est facile d’identifier la présence de l’insecte par le long filament cireux que celui-ci sécrète à l’extérieur du tronc donnant l’apparence d’un poil ou d’un cheveu (photo 3). Les dommages causés par la cochenille filamenteuse rendent l’écorce du hêtre plus rugueuse, créant ainsi des sites favorables à l’établissement de la cochenille du hêtre et à l’introduction des champignons.

Photo 2 – Dégâts caractéristiques de la cochenille filamenteuse (photo Sébastien Meunier).

Photo 3 – Filament cireux secrété par la cochenille filamenteuse (photo N. Nadeau-Thibodeau).
Cycle biologique de la maladie et symptômes

Photo 4 – Taches brunâtres sur l’écorce du hêtre à la suite de l’infection par Neonectria (Photo : Sébastien Meunier)
Les spores du champignon (ascospores) sont disséminées par la pluie et le vent et se logent dans les microblessures pour infecter le cambium. On peut alors observer des taches brunâtres sur l’écorce (photo 4), puis graduellement, la nécrose du cambium suivie d’un affaissement localisé de l’écorce. C’est alors que l’on remarque l’apparition de petits chancres circulaires d’environ 2 cm de diamètre (photo 5). Ceux-ci peuvent se fusionner lorsque l’arbre est gravement atteint donnant au tronc un aspect verruqueux. Par la suite, on observe un dépérissement du houppier, un jaunissement du feuillage et la mort survient à brève échéance (photo 6). Les hêtres de plus fort diamètre, ayant une écorce plus rugueuse et étant couverts de cochenilles du hêtre sont les plus sensibles à la maladie.

Photo 5 – Petits chancres circulaires (photo : N. Nadeau-Thibodeau)

Photo 6 – Dépérissement du houppier des hêtres à grandes feuilles (photo MFFP).
Les fructifications du champignon, les périthèces, se développent sur l’écorce trois à cinq ans après l’invasion de la cochenille (photo 7). Elles apparaissent sous forme de petites boules rouge orangé. Elles atteignent leur maturité vers la fin de l’été (photo 8). Ainsi, les périthèces produiront l’année suivante des ascospores à l’origine de nouvelles infections. Les champignons possèdent aussi une forme anamorphe (asexuée) appelée Cylindrocarpon faginatum C.Booth.et C. heteronema (Berk. & Broome) Wollenw. Ils produisent des macroconidies qui peuvent être dispersées du mois de juillet jusqu’à la fin de l’automne.

Photo 7 – Tronc de hêtre couvert de périthèces (photo N. Nadeau-Thibodeau)

Photo 8 – Périthèces de Neonectria à maturité (photo N. Nadeau-Thibodeau)
Évolution de la maladie et distribution
Évolution de la maladie corticale du hêtre au Québec 2019
Dégradation des bois et impacts
Ressource ligneuse
Le processus de dégradation du bois de hêtre est assez rapide, mais il diffère légèrement selon la source du problème (chablis partiel, MCH, blessures d’exploitation, sénescence naturelle). Le bois de hêtre est très sensible à la carie de l’aubier et du cœur, quelle que soit l’origine de l’infection. La vitesse de dégradation du bois chez les arbres atteint de la MCH varie selon leur diamètre : les plus gros sont plus résistants que ceux de 30 cm ou moins de diamètre qui, eux, cassent les premiers, souvent avant même de mourir sur pied.
- Les hêtres de 30 cm ou moins de diamètre
Ces arbres malades ont généralement le cœur sain. Les chancres diffus sur une bonne partie du tronc entraînent avec le temps le dépérissement du houppier. Le débitage de hêtres malades montre qu’ils peuvent survivre plus de 20 ans sans montrer le moindre symptôme de dépérissement. Sitôt qu’il y a plus de 10 % de feuillage dépéri du houppier, l’arbre est par contre condamné à brève échéance. Les hêtres survivent rarement plus de cinq ans et le processus de dégradation de l’aubier s’amorce bien avant que l’arbre ne meure. Le sylviculteur dispose d’un délai de deux ans tout au plus pour récupérer du bon bois de chauffage provenant de hêtres dont le houppier a commencé à dépérir. Le feuillage est alors clairsemé et chlorotique, et le nanisme foliaire touche le houppier. Ces arbres meurent sur pied ou cassent sous le houppier, victimes de la carie de l’aubier causée surtout par les champignons Trichaptum pargamenum (Fr.) G. Cunn., Trametes versicolor (L.:Fr.) Pilát ou Fomes fomentarius (L.:Fr.) Kickx. Le plus souvent, les arbres cassent avant même que les champignons n’apparaissent sur le tronc.
- Les hêtres de 30 cm ou plus de diamètre
Les gros sujets dont le houppier est encore vigoureux survivent plus longtemps, parfois plus d’une période de rotation. Ceux qui meurent sur pied sont habituellement victimes du pourridié-agaric (Armillaria ostoyae (Romagn.) Herink) qui avait déjà envahi les racines au cours de la décennie précédant l’infection par la MCH. Avec le temps, les grands hêtres malades voient leur écorce décoller par endroits, montrant alors des symptômes de carie de l’aubier. Ceux atteints en profondeur affichent des sporophores (par exemple : Fomes fomentarius (L.:Fr.) Kickx, Ganoderma applanatum (Pers.) Pat., Trametes versicolor (L.:Fr.) Pilát, Trichaptum pargamenum (Fr.) G. Cunn). Contrairement à Phellinus igniarius (L.:Fr.) Quél., qui dégrade le bois de cœur, tous les autres champignons causent une carie de l’aubier bien avant que les arbres ne meurent et même les plus gros peuvent casser par grand vent. La carie s’étend rapidement dans l’aubier des hêtres malades et si ces derniers sont déjà atteints au cœur, ils sont généralement irrécupérables comme bois d’œuvre. Il est donc recommandé de récupérer en priorité les hêtres malades bien avant que les sporophores n’apparaissent sur le tronc.
Ce que nous savons du processus de dégradation du hêtre dans l’est (Bas-Saint-Laurent, Gaspésie-Îles-de-la-Madeleine) ne signifie pas que ce soit la même chose qui se produira dans les régions de l’Ouest (Outaouais, Abitibi-Témiscamingue) au cours des prochaines décennies. Comme le climat est plus sec et plus chaud et que la saison de croissance des arbres (et des champignons) est plus longue, le pire est à prévoir. La maladie corticale risque fort de se développer plus rapidement et de tuer les hêtres en grand nombre et sur une plus courte période, compte tenu de l’interaction insidieuse du pourridié-agaric qui se manifeste surtout après de longs épisodes de sécheresse. De plus, la carie des racines risque de causer des chablis inhabituels lors d’ouragans ou tornades dont la force et la fréquence sont plus grandes dans l’Ouest que dans l’Est de la province. La phase de dévastation de la MCH risque fort de survenir avant 20 ans, compte tenu de la forte proportion de grands hêtres sénescents qui seront infectés par la maladie corticale. Les érablières qui ont déjà été exploitées en coupe partielle par le passé renferment en effet une plus forte proportion de hêtres parvenus à maturité.
Faune
La MCH cause aussi de nombreux impacts sur la faune. La dégradation des hêtres touche entre autres l’habitat et la disponibilité de nourriture de plusieurs espèces animales. Par exemple, les populations de grands pics profitent avantageusement des effets de la MCH puisque les hêtres malades constituent un refuge de choix pour la nidification et une source de nourriture abondante d’insectes xylophages qui y prolifèrent. Les effets sont par contre différents chez les espèces se nourrissant des faînes de hêtre. D’après une étude effectuée dans le domaine bioclimatique de l’Érablière à bouleau jaune dans l’ouest du Québec (Bergeron et Dumont 2000), la biomasse de faînes de hêtre produite atteint dans certaines strates forestière 103 kg/ha. Ces semences sont très recherchées par de nombreux rongeurs, certaines espèces d’oiseaux (dont le dindon sauvage, Meleagris gallopavo, et la gélinotte huppée, Bonasa umbellus), le cerf de Virginie, Odocoileus virginianus, et l’ours noir, Ursus americanus. Le hêtre est à ce point recherché par les ours noirs que ces derniers grimpent aux arbres pour consommer les faînes avant leur chute au sol. La mortalité des hêtres de gros diamètre cumulée à la perte de vigueur des arbres affectés par la MCH entraînent une diminution de la production de faînes. Selon la littérature, il y aurait une relation de cause à effet entre la production de faînes et les populations d’ours noirs. Les faînes sont une source importante de nourriture surtout pendant la période précédant l’hibernation. Le succès de reproduction des ours noirs dépendrait du volume de faînes disponibles (Jakubas et coll. 2004). Les chercheurs ont observé que le taux de femelles fécondes n’était que de 22 % après une mauvaise année semencière, comparativement à 80 % après une forte production de faînes l’automne précédent.
Sécurité publique
La mortalité des hêtres associée à cette maladie représente un problème de sécurité publique en raison de la présence de nombreux hêtres dans les parcs urbains, les terrains de camping ainsi qu’en bordure des routes et des sentiers de villégiature. Le cas vécu en septembre 2012 au Killbear Provincial Park en Ontario  nous rappelle que ce problème n’est pas qu’un enjeu d’approvisionnement ligneux, mais qu’il est aussi un enjeu de sécurité publique. Les autorités du parc ont dû faire abattre des centaines de hêtres malades qui menaçaient de rompre à tout moment. Les arbres malades deviennent rapidement une menace sérieuse pour la sécurité des personnes et des biens, surtout dans les sites de villégiature.
nous rappelle que ce problème n’est pas qu’un enjeu d’approvisionnement ligneux, mais qu’il est aussi un enjeu de sécurité publique. Les autorités du parc ont dû faire abattre des centaines de hêtres malades qui menaçaient de rompre à tout moment. Les arbres malades deviennent rapidement une menace sérieuse pour la sécurité des personnes et des biens, surtout dans les sites de villégiature.
Impact sur la dynamique forestière
Envahissement par les drageons de hêtre
Un autre problème bien connu dans le monde forestier est l’envahissement des érablières par le hêtre. Plusieurs hypothèses ont été avancées pour expliquer cet envahissement, entre autres l’effet indirect de la MCH. Les hêtres malades sont d’abord enclins à produire de nombreux drageons. Les ouvertures du couvert engendrées par la mort des grands hêtres apportent par la suite des conditions de lumière accrues favorables à la croissance en hauteur des drageons de hêtres préétablis dans les étages inférieurs du couvert.
Recommandations d’aménagement
Il n’existe pas de solution efficace pour réduire les sources d’inocula d’agents pathogènes ni de moyens directs pour maîtriser les populations de cochenilles. Cependant, certaines interventions contribuent à réduire la vulnérabilité des peuplements, tout en limitant l’interférence de certaines espèces comme le hêtre, l’érable de Pennsylvanie, Acer pensylvanicum L., et la viorne, Viburnum, sur la régénération en essences feuillues recherchées. Le hêtre demeure exploitable dans la mesure où les tiges renferment du bois d’œuvre ou du bois destiné aux pâtes et papiers.
Un atelier de travail sur l’aménagement des érablières dont les hêtres sont affectés par la maladie corticale a eu lieu les 28, 29 et 30 septembre 2015 à Huntsville en Ontario. Des réflexions découlant de cet atelier, il a été convenu de produire un guide des meilleures pratiques d’aménagement pour ces érablières.
Dans l’attente de la parution du guide au cours de 2016, nous formulons des recommandations s’inspirant de travaux antérieurs (McCullough et coll. 2005) et du cadre d’intervention adopté en Ontario qui est fondé sur les trois phases d’évolution de la maladie (McLaughlin et Greifenhagen 2012).
Peuplement sain sans signe ni symptôme de maladie dans toutes les zones de déploiement
Un tel peuplement peut se situer dans la zone d’invasion ou dans une autre zone s’il a été jusqu’ici épargné. Les peuplements contenant plus de 25 % de la surface terrière (ST) totale en hêtres sont les plus vulnérables à la MCH. Par contre, les dommages appréhendés sont d’autant plus faibles que le volume de hêtres est faible dans le peuplement. On pourra alors simplement gérer l’infestation avec la méthode du jardinage. Toutefois, si la haute régénération est dominée par le hêtre, un dégagement mécanique des érables peut s’avérer nécessaire.
- Cas 1 : composante mineure de hêtre (ST < 25 %)
L’impact appréhendé est de faible importance. Le marteleur pourra reconnaître facilement et protéger les hêtres semenciers convoités par l’ours noir. Le couvert peut encore être maintenu même avec la disparition des hêtres infectés.
- Cas 2 : composante majeure de hêtre (ST > 25 %)
L’impact appréhendé est important. Il faut réduire la présence du hêtre et repérer les secteurs mal régénérés en érable à sucre ou dominés en sous-étage par des espèces comme le hêtre, la viorne et l’érable de Pennsylvanie. La création de trouées serait souhaitable dans ces secteurs particuliers afin de favoriser le recrutement en essences feuillues recherchées. Dans les cas de peuplements qui ont été convertis en hêtraie pure ou en érablière rouge à hêtre, une approche par coupes progressives pourrait s’appliquer. L’abondance de semis et de drageons de hêtre avant la récolte peut aider à moduler le traitement afin de s’assurer d’un retour des essences feuillues telles que le bouleau jaune, Betula alleghaniensis Britt., le tilleul d’Amérique, Tilia americana L., le chêne rouge, Quercus rubra L., et l’érable à sucre, Acer saccharum Marsh. Sur une petite échelle, il faut au préalable réduire la densité de la strate de gaules et de perches du hêtre afin d’aider les autres essences à s’établir avant la coupe de l’étage supérieur.
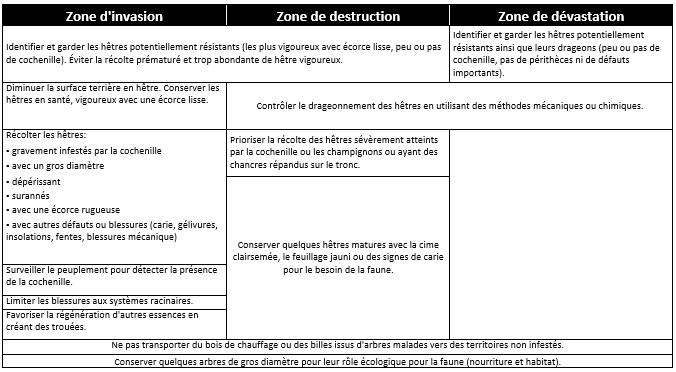
En zone d’invasion
Il est recommandé de marquer pour la coupe les arbres montrant un feuillage chlorotique ou un dépérissement au niveau du houppier. Il faut aussi marquer les arbres gravement infestés par l’insecte. Les arbres peu infestés sont encore considérés comme étant résistants à la maladie. Au Nouveau-Brunswick, on aurait observé un taux moyen de 3,3 % de hêtres résistants (Taylor et al. 2013) pour la région de la forêt acadienne alors que d’autres sources font état d’un taux de résistance de l’ordre de 1 %. On doit donc favoriser la croissance des tiges appartenant à l’étage inférieur du couvert et relativement peu infestées par les cochenilles.
En zone de destruction ou de dévastation
Il est prioritaire de couper les arbres montrant des lésions et des nécroses du cambium, car ces arbres comportent des défauts importants qui réduisent le potentiel de sciage. Il faut par contre protéger les arbres affichant une résistance partielle à la maladie. Ces derniers sont faciles à repérer, car ils forment des groupes d’arbres issus d’un même clone. Il faut du même coup assainir le peuplement en récoltant les arbres moribonds. Les arbres morts vont graduellement être renversés par le vent ou se rompre pour générer des chicots constituant des refuges de choix pour de nombreuses espèces animales (pics, polatouches, etc.). En se décomposant, les troncs renversés fourniront un terreau fertile à l’établissement d’essences en raréfaction, comme le bouleau jaune, l’épinette rouge, Picea rubens Sarg., le thuya occidental, Thuya occidentalis L., et la pruche du Canada, Tsuga canadensis (L.) Carr.
Le problème de recrutement d’essences nobles peut nécessiter un travail d’élimination des gaules de hêtre, avant d’avoir effectué, ou après, la récolte d’arbres par différentes coupes partielles. Il convient d’accorder une importance prioritaire à la sécurité des travailleurs forestiers en repérant et en marquant les arbres dangereux tel qu’il est indiqué dans le guide de reconnaissance des défauts des arbres (Boulet et Landry 2015).
Le tableau ci-dessous présente une synthèse des recommandations d’aménagement décrites précédemment.
Cliquez pour agrandir
Références
Bergeron, N. et A. Dumont, 2000, Mesure de la biomasse de faînes de hêtres à grandes feuilles (Fagus grandifolia) et de glands de chênes rouge (Quercus rubra) produite par les forêts feuillues matures du domaine bioclimatique de l’Érablière à bouleau jaune dans l’ouest du Québec. Université du Québec à Rimouski, Département de biologie, 32 pages + cartes.
Boulet, B., et G. Landry. 2015. La carie des arbres – Fondements, diagnostic et application. 3e édition, Les Publications du Québec, Québec. 340 p.
Jakubas, W.J., C. R. McLaughlin et P. G. Jensen. 2004. Alternate year beechnut production and its influence on bear and marten population. Proc. Beech Bark Disease Symposium, pp. 79-87. USDA For. Serv. Gen. Tech. Rep. NE-331.
McCullough, D.G., R.L. Heyd et J.G. O’Brien 2005. Biology and management of beech bark disease : Michigan’s newest exotic forest pest. Michigan State University Extension, bulletin E-2746.
McLaughlin J., et S. Greifenhagen. 2012. Beech bark disease in Ontario: a primer and management recommendations. Ontario Forest Research Institute, Forest research note no. 71.
Ministère des Ressources naturelles. 2013. Le Guide sylvicole du Québec, Tome 1, Les fondements biologiques de la sylviculture, ouvrage collectif sous la supervision de B. Boulet et M. Huot, Les Publications du Québec, 1044 p.
Roy, M.-E., et P. Nolet. 2015. Typologie et caractérisation spatiale de l’envahissement et de la maladie corticale du hêtre en Outaouais. Institut des sciences de la forêt tempérée. Université du Québec en Outaouais. Ripon, Québec. Rapport technique. 62 p. + Annexes.
Taylor, A. R., D. A. McPhee et J. A. Loo. 2013. Incidence of beech bark disease resistance in the eastern Acadian forest of North America. The Forestry Chronicle vol. 89, no. 5, p. 690-695.
